Director and Founder of SCP, Kevin Hunt
Kevin D. Hunt was born in Indianapolis, Indiana and moved with his family to Oak Ridge, Tennessee just before he reached his teens. In high school he took an interest in human evolution, and matriculated to the University of Tennessee to study Human Paleontology under Fred Smith (B.A., 1980). He continued at the University of Michigan (M.A., 1982; PhD, 1989) where his doctoral work was supervised by C. Loring Brace and Richard W. Wrangham. During his graduate years he worked several years in Brace’s lab crunching numbers for several publications on dental evolution among Neanderthals and early modern humans. He collaborated with Brace in investigating patterns of human dental and cranial variation, most significantly in "A Non-Racial Craniofacial Perspective on Human Variation: A(ustralia) to Z(uni)" (Am. J. Phys. Anthropol., 1990) among other publications.
For his doctoral research he documented locomotion and posture (positional behavior) among chimpanzees and baboons in 1986 and 1987. He shuttled back and forth between the Mahale Mountains National Park, where he was affiliated with Toshisada Nishida’s research project, and Gombe National Park, where he shared a few chimpanzee follows with Jane Goodall herself.
 Hunt was a Postdoctoral Fellow at Harvard University from 1989 to 1991 where he worked with Richard Wrangham as part of Wrangham’s study of primate feeding behavior at the Kibale Forest Reserve, Uganda. He studied the feeding behavior of chimpanzees, red colobus monkeys, black and white colobus monkeys, mangabeys, redtail monkeys and blue monkeys over three stints in the Kibale Forest. One six month visit he was accompanied by his wife and their two year old twins. Among the publications arising from this collaboration were “The significance of fibrous food for Kibale Forest chimpanzees (Philosophical Transactions of the Royal Society (B) 334: 171-178) and “Dietary response of chimpanzees and cercopithecines to seasonal variation in fruit abundance. I. Antifeedants” (International Journal of Primatology. 19: 949-970).
Hunt was a Postdoctoral Fellow at Harvard University from 1989 to 1991 where he worked with Richard Wrangham as part of Wrangham’s study of primate feeding behavior at the Kibale Forest Reserve, Uganda. He studied the feeding behavior of chimpanzees, red colobus monkeys, black and white colobus monkeys, mangabeys, redtail monkeys and blue monkeys over three stints in the Kibale Forest. One six month visit he was accompanied by his wife and their two year old twins. Among the publications arising from this collaboration were “The significance of fibrous food for Kibale Forest chimpanzees (Philosophical Transactions of the Royal Society (B) 334: 171-178) and “Dietary response of chimpanzees and cercopithecines to seasonal variation in fruit abundance. I. Antifeedants” (International Journal of Primatology. 19: 949-970).
 Several researchers suggested that bipedalism was an adaptation for gathering of food from grasses, trees or bushes (DuBrul, 1962; Jolly, 1970; Rose, 1976, 1984, 1991; Wrangham, 1980). The seed eating hypothesis in particular (Jolly, 1970) focused on the demands of collecting small, evenly distributed resources close to ground level. It quickly became obvious that primates simply pull the grass over when it is too tall to reach. The small-object feeding hypothesis became a more widely accepted refinement of the seed-eater model; it substituted small fruits for seeds (Rose, 1976; Jolly and White, 1995). Wrangham (1980) offered a locomotor corollary to the small object feeding hypothesis, arguing that bipedal locomotion saves energy by eliminating the action of repeatedly raising the upper body to feed bipedally after walking between resources in a typical primate quadrupedal fashion (Jolly, 1970; ; Jolly & White, 1995). Terrestrial feeding hypotheses are straightforwardly synthesized into a Jolly/Rose/Wrangham hypothesis that postulates bipedal posture evolved because it was advantageous for reaching up into trees, and that bipedal locomotion evolved to reduce energy costs when traveling between densely packed feeding sites.
Several researchers suggested that bipedalism was an adaptation for gathering of food from grasses, trees or bushes (DuBrul, 1962; Jolly, 1970; Rose, 1976, 1984, 1991; Wrangham, 1980). The seed eating hypothesis in particular (Jolly, 1970) focused on the demands of collecting small, evenly distributed resources close to ground level. It quickly became obvious that primates simply pull the grass over when it is too tall to reach. The small-object feeding hypothesis became a more widely accepted refinement of the seed-eater model; it substituted small fruits for seeds (Rose, 1976; Jolly and White, 1995). Wrangham (1980) offered a locomotor corollary to the small object feeding hypothesis, arguing that bipedal locomotion saves energy by eliminating the action of repeatedly raising the upper body to feed bipedally after walking between resources in a typical primate quadrupedal fashion (Jolly, 1970; ; Jolly & White, 1995). Terrestrial feeding hypotheses are straightforwardly synthesized into a Jolly/Rose/Wrangham hypothesis that postulates bipedal posture evolved because it was advantageous for reaching up into trees, and that bipedal locomotion evolved to reduce energy costs when traveling between densely packed feeding sites.
Hunt observed that while chimpanzees did feed bipedally not infrequently, but they did so not simply by reaching up into the trees from the ground. Instead, chimpanzee bipedal feeding was half in arboreal and half in terrestrial contexts (Hunt, 1994a). The food items on which chimpanzees fed bipedally were distinctive. Most were fruits which were quite small (1-5 mm) and were found in short trees (1-15 m). When reaching up from the ground, small fruits encourage bipedalism because it takes the collecting rate of both hands to keep the mouth full, whereas large fruits may be plucked off and chewed while sitting. Small food items require finer manipulation to harvest, compared to the larger food items more typical of a chimpanzee diet.
After his postdoc, Hunt was hired at Indiana University, where he continued to synthesize work on ape ecology, ape anatomy and early human evolution.
 Australopith morphology from the lumbar vertebrae caudad is an amalgam of ape-like features, human-like features, and unique features, all consistent with a postural feeding hypothesis. The human-like pelves of A. afarensis (Lovejoy 1988) and A. africanus (Abitbol 1995, Robinson 1972) indicate habitual bipedalism. Cross-sectional properties of the A. afarensis humerus and femur resemble modern humans more than apes (Ohman, 1993; Ohman et al., 1997). Other features which are less obvious, such as long curved toes, less stable knee-joints, wide hips, and short hindlimbs suggest this bipedalism was functionally different from that of modern humans. Hunt interprets the wide pelvis of australopiths—much wider than necessary for parturition, and resembling neither apes nor humans—as evolved to lower the center of gravity to improve balance on arboreal supports (Hunt 1994a, 1996, 1998). Such a broad pelvis suggests that energy-conserving and low-stress bipedal locomotion did not evolve until Homo erectus. The short australopith lower limb (Jungers 1982, 1991) is a corollary adaptation for lowering the center of gravity (Hunt, 1996). The previously unexplained hyper-valgus femur of australopiths is not an independent adaptation, or perhaps not even under direct selection (Tardieu et al. 2006), but rather a mechanical response to a short femur and wide pelvis.
Australopith morphology from the lumbar vertebrae caudad is an amalgam of ape-like features, human-like features, and unique features, all consistent with a postural feeding hypothesis. The human-like pelves of A. afarensis (Lovejoy 1988) and A. africanus (Abitbol 1995, Robinson 1972) indicate habitual bipedalism. Cross-sectional properties of the A. afarensis humerus and femur resemble modern humans more than apes (Ohman, 1993; Ohman et al., 1997). Other features which are less obvious, such as long curved toes, less stable knee-joints, wide hips, and short hindlimbs suggest this bipedalism was functionally different from that of modern humans. Hunt interprets the wide pelvis of australopiths—much wider than necessary for parturition, and resembling neither apes nor humans—as evolved to lower the center of gravity to improve balance on arboreal supports (Hunt 1994a, 1996, 1998). Such a broad pelvis suggests that energy-conserving and low-stress bipedal locomotion did not evolve until Homo erectus. The short australopith lower limb (Jungers 1982, 1991) is a corollary adaptation for lowering the center of gravity (Hunt, 1996). The previously unexplained hyper-valgus femur of australopiths is not an independent adaptation, or perhaps not even under direct selection (Tardieu et al. 2006), but rather a mechanical response to a short femur and wide pelvis.
An antero-posteriorly short, rounded lateral femoral condyle (for some specimens, Tardieu 1983, McHenry 1986) and a convex lateral tibial condyle (pers. obs.) suggest great flexibility of the lower limb, compromising knee stability and thus limiting long-distance walking.
While the australopith foot is strut-like and shock-absorbing (Latimer & Lovejoy 1990, Langdon et al. 1991) with a modern large calcaneus and robust hallux, other pedal and leg anatomy is ape-like. The medial cuneiform of A. afarensis is convex, suggesting at least a rudimentary first toe gripping ability (Tuttle 1981, Stern & Susman 1983, Deloison 1991), pedal phalanges, including the hallux, are long, curved (Tuttle 1981, Stern & Susman 1983) and have plantar-oriented metatarsophalangeal joint surfaces (Duncan et al. 1994), giving a gripping set to the toe joints. 
The absence of an abductible great toe and the retention of long, curved, gripping toes may seem contradictory at first glance. The in-line great toe has been interpreted by some specialists to be incontrovertible evidence that australopiths were obligate terrestrial walkers, and long gripping toes is seen as merely a primitive retention. Chimpanzee behavior suggests that a gripping great toe is unnecessary for climbing short trees, but lateral toes are used to grip small branches (Hunt 1998).
This suggests that chimpanzees confined to open habitats where many of their resources are found in shorter trees, we might find behavior and perhaps morphology (though any true specializations might be swamped by breeding with nearby forest-adapted chimpanzees). Prof. Hunt initiated research at Semliki to test this hypothesis. In data gathered so far at Semliki, though not yet published, it is clear that the chimpanzees engage in enough bipedal behavior that they have anatomical adaptations that make them more human-like than other chimpanzees.
 Because Semliki chimpanzees feed among the abundant short trees there, they are a perfect population on which to test these hypothesis.
Because Semliki chimpanzees feed among the abundant short trees there, they are a perfect population on which to test these hypothesis.
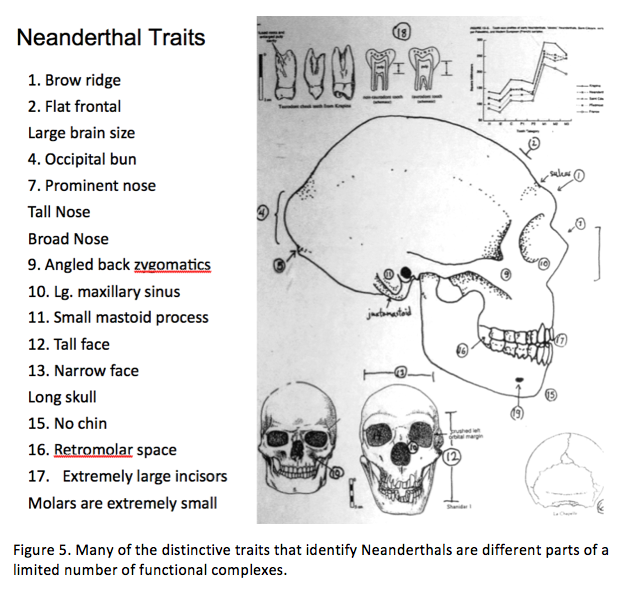
While he continues this research, he also has worked on primate fossils and Neanderthal adaptations. His work suggests that Neanderthal faces in particular are much less specialized than many maintain
Prof. Hunt lives in Bloomington, IN with his wife Marion Gewartowski Hunt and their three children Alison, Daniel and David. He was pleased to share Semliki with his daughter in 2010, the first time Alison had returned to Uganda since 1993. In his spare time he enjoys hiking, cycling, basketball and tennis. A complete Curriculum Vita can be found here.

{kind=link}
{kind=link}
{kind=link}
{kind=link}